|
||||
|
|
Глава VIII ГОЛОГРАММЫ ГИПОТЕЗА Вернемся на время к нашей общей модели. Модель предполагает два основных процесса: пространственно организованные-состояния и операции, выполняемые на фоне этих состояний с помощью импульсной передачи между нейронами. В первой части книги было описано, каким образом эти основные свойства нейронных групп могут комбинироваться в логические операции, усиливающие аналитические и контрольные (серво) функции" нервной системы. Учитывая их значение, а также то, что сейчаа они игнорируются в нейропсихологической и нейрофизиологической литературе, я подчеркнул, что построение имеющих определенную структуру топологических, то есть пространственных, представительств в нервной системе является одной из форм, которые могут принимать состояния мозга. Я предположил, что-взаимодействие динамических структур возбуждения, падающих на рецепторные поверхности, после их передачи по параллельным путям кодируется благодаря горизонтальным связям в активность медленных потенциалов групп нейронов и образует временные микроструктуры, рисунки которых зависят скорее от функциональной организации нейронных соединений, чем от нейронов, как таковых. В гл. VII были подробно рассмотрены факты, свидетельствующие о существовании детекции признаков и механизма анализа, а также приведены аргументы в пользу необходимости особой нейронной организации, помимо той, которая представлена анализаторами признаков. В гл. VIII формулируется гипотеза о том, что такой организацией может быть механизм микроструктуры медленных потенциалов соединительных аппаратов. Эта гипотеза основывается на предположении, что нейронное отображение входных воздействий не является фотографическим и создается не только посредством имеющейся системы фильтров, выделяющих признаки, но и с помощью особого класса преобразований, которые обладают значительным формальным сходством с процессом отражения оптического образа, открытым математиками и инженерами. Этот оптический процесс, названный голографией, основан на использовании явления интерференции структур. Он обладает множеством удивительных свойств, из которых первостепенное значение имеет его способность к распределению и сохранению большого количества информации. Именно эти свойства дают возможность разрешить противоречие между потребностью в функциональной лабильности, быстром темпе изменений и уже рассмотренными анатомическими особенностями в организации нервных систем приемл информации. Прежде чем точно сформулировать проблему, полезно посвятить несколько параграфов разъяснению нашего общего подхода. Обработка оптической информации с помощью голографии описывается математически в понятиях механической волны. В физической оптике для описания распространения света в экспериментальных условиях могут быть использованы как квантовые, так и волновые уравнения. Физика не интересует то, является ли свет квантом или волной, распространяется ли он в виде частиц или электромагнитных волн, или в форме того и другого. Он заботится о том, чтобы как можно полнее количественно описать результаты своих наблюдений, и соответственно этому выбирает инструменты для своего описания. Некоторые наблюдения могут быть хорошо описаны как статистическая вероятность квантовых событий, другие легче описываются с помощью волновой теории. Особенно трудно привести в соответствие с квантовым подходом одно наблюдение. Когда свет очень низкой интенсивности проходит через решетку, то есть когда предполагается, что через каждую отдельную щель пройдет самое большее несколько квантов света, уравнение, описывающее световой поток по другую сторону решетки, для объяснения наблюдаемых эффектов должно учитывать некоторое взаимодействие, которое, по-видимому, имеет место между световыми потоками, проходящими через соседние щели (рис. VIII-1). 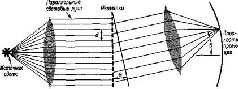
Трудно, хотя и не невозможно, объяснить это взаимодействие с точки зрения квантовой теории: каким образом квант света может влиять на соседние, отделенные от него решеткой? Кажется, будто каждый квант оказывает силовое (возникает искушение сказать «магнетическое») действие на соседний. Этот феномен взаимодействия был широко изучен и получил название суперпозиции. Математические уравнения, удовлетворительно объясняющие явление суперпозиции, представляют собой систему линейных уравнений, названных интегралами свертки, которые обычно используются для описания взаимодействия волновых процессов. Описание одной волновой формы свертывается вместе с описанием другой. При объяснении суперпозиции не возникает серьезных теоретических проблем, если считать, что ровный фронт волн света, встречаясь с решеткой, разбивается ею на небольшие, взаимодействующие между собой «вихри». Можно считать, что такие вихри и вызывают эффект интерференции и объясняют суперпозицию – «взаимодействие по соседству». Голографическая гипотеза функционирования мозга в процессе восприятия принимает форму суперпозиции. Фактически выбор способа объяснения того, как изменяются электрические потенциалы мозга – в статистических терминах или в волновых,- зависит от наблюдений, которые должны быть описаны. Следует выбрать тот способ, который может дать наиболее полное количественное описание наблюдений и в то же время быть приемлемым для концепции. В гл. III описано возникновение латерального, или периферического, торможения в рецепторах и на разных уровнях центральной нервной системы. Латеральное торможение обусловливает эффект взаимодействия между процессами, протекающими в соседних нейронах. Если это пространственное взаимодействие адекватно описывается интегралами свертки, которые характеризуют физическое явление суперпозиции, то мы можем рассматривать взаимодействие в нервной системе на основе интерференционных эффектов. Правомочность такого выбора подтверждается теми фактами, что по крайней мере в сетчатке взаимодействие между соседними горизонтальными клетками обусловлено исключительно тормозными взаимодействиями посредством медленных потенциалов гиперполяризации, а не деполяризации, которая приводит к возникновению нервных импульсов. Как и в гл. III, рецепторные явления служат миниатюрными моделями на этот раз «нейронного голографического» процесса. Мы уже отмечали, что возбуждение одной единицы зрительного нерва оказывает влияние на частоту разрядов соседних единиц. Мы отметили также, что рецептивное поле отдельной единицы образуется в результате такого пространственного взаимодействия между соседними элементами. В зрительном нерве эти рецептивные поля обычно состоят из более или менее округлого центрального пятна, которое реагирует либо возрастанием (on – Центральный нейрон), либо снижением (off – центральный нейрон) частоты своих спонтанных разрядов, и из окружающей зоны, которая характеризуется активностью, противоположной по знаку активности центра. Глава VII была посвящена проблеме: .какой тип кода, какой тип организации создается в результате действия многих таких единиц? Настоящая глава посвящена другому кругу проблем, а именно: что кодируется в одном рецептивном поле, то есть какие черты оптического изображения определяют форму рецептивных полей? Ответ на этот вопрос следует из проделанного Родие-ком (1965) детального математического анализа количественных отношений, существующих между специфическим образом движущимися зрительными стимулами (вспышками) и соответствующими им формами рецептивного поля. Кривые полученных ответов (каждая из которых представляет собой вертикальный срез через трехмерное рецептивное поле, рис. VIII-2) имеют особую форму. При одновременном предъявлении двух или более стимулов кривая ответов достаточно хорошо соответствует кривой, которая возникает в результате суперпозиции кривых ответов на раздельные предъявления этих стимулов. Таким образом, Родиек делает зывод, что, исходя «из структур ответа на небольшое световое пятно, возникающих при засвечивании разных участков рецептивного поля клетки, можно получить структуру ответа этой клетки на фигуру любой формы, движущуюся в любом направлении, с любой скоростью, через любую часть рецептивного поля». Это открытие позволило воспроизвести рецептивное поле клетки, используя только движущиеся пятна света, а также сделало возможным с помощью компьютера произвести запись, показанную на рис. VIII-7. Таким образом, форма рецептивного поля оказывается сверткой первой производной от формы зрительного стимула. Как уже отмечалось, интегралы свертки, устанавливающие соотношения между событиями, происходящими в соседних областях (пространственная суперпозиция), описывают основной го-лографический процесс в системах, перерабатывающих оптическую информацию (Gabor, 1949, 1951). Эффекты интерференции описываются также уравнениями преобразований Фурье, Френеля и Лапласа. Определение же того, какое преобразование больше подходит для описания действия системы в данных условиях, является эмпирической проблемой. 
В зрительной системе – и имеются достаточные основания благодаря работам Г. фон Бекеши (1960, 1967, рис. VIII-3), чтобы предполагать, что также и в слуховой, соматической и вкусовой системах, – эти виды преобразований с большой точностью описывают взаимодействие между структурами энергетических воздействий, возбуждающими рецепторные поверхности, ж спонтанными изменениями потенциалов в рецепторных единицах. В системах, обрабатывающих оптическую информацию, такие преобразования обычно имеют отношение к анализу формы волны (то есть эффектов интерференции), поэтому нет особых причин полностью игнорировать волновой подход и при объяснении эффекта суперпозиции, возникающего в нейронных системах. Преимущество такого подхода состоит в том, что физический процесс, основывающийся на эффектах интерференции, обладает многими свойствами нервного процесса восприятия и, таким образом, облегчает понимание мозгового механизма формирования образов. Рассмотрим теперь физическую голограмму. 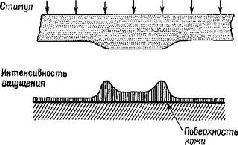
ФИЗИЧЕСКАЯ ГОЛОГРАММА Большинству из нас известно, каким образом возникает изображение в обычных оптических системах. Фотоаппарат регистрирует на фотопленке, помещенной в плоскости изображения, копию пространственного распределения интенсивности света, отраженного от предметов, попавших в поле объектива. Каждая точка на фотопленке сохраняет информацию, полученную от соответствующей точки зрительного поля, и, таким образом, зарегистрированная картина выглядит подобно картине зрительного поля. Недавно-были проведены исследования свойств записей, сделанных на фотопленке, которая находится в оптической системе где-то перед плоскостью изображения (то есть в фокальной плоскости). При соответствующей экспозиции такой фотопленки с помощью источника когерентного света она становится оптическим фильтром, в котором информация о каждой точке зрительного поля распределяется и хранится по всему фильтру (рис. VIII-4). 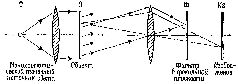
Такие фильтры обладают многими удивительными свойствами. Как мы знаем, когда фотопленка внутри камеры находится не точно в плоскости оптического изображения, образ становится расплывчатым, его границы менее четки, контраст менее выражен. В оптическом фильтре информация распределяется таким образом, что зафиксированное на пленке изображение вообще не является подобием зрительного изображения. Оптический фильтр представляет собой запись волновых узоров, излучаемых объектом или отражаемых от него. «Такую запись можно рассматривать как «замороженный» волновой узор, который остается «замороженным» до тех пор, пока не приходит время восстановить процесс, после чего волны «считываются» с регистрирующего посредника» (Leith and Upatniecks, 1965). Так, когда оптический фильтр освещается источником когерентного света, он восстанавливает волновые фронты света, которые имели место при экспозиции объекта. В результате при рассматривании фильтра становится видно действительное изображение зрительного поля. Этот зрительный образ, по-видимому, точно совпадает с картиной, имевшей место при экспозиции, он является ее полным трехмерным изображением (рис. VIII-5). По существу, вся информация, описывающая зрительное поле, из которой может быть восстановлено изображение, содержится в этом фильтре. 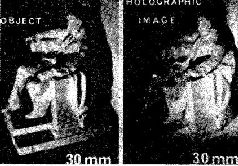
«Когда наблюдатель при рассмотрении фильтра меняет свое положение, перспектива изображения тоже меняется, как если бы наблюдатель рассматривал реальную сцену. Обнаружены эффекты параллакса между близко и далеко расположенными объектами: если один предмет находится на переднем плане и заслоняет какой-то другой предмет, то наблюдатель может, двигая головой, заглянуть за заслоняющий предмет и: таким образом увидеть то, что за ним скрывается… Короче говоря, восстановленная голограмма обладает всеми визуальными свойствами реальной сцены, и мы не знаем ни одного зрительного теста, который можно было бы использовать, чтобы отличить их друг от друга (Leith and Upatnieeks,. 1965, p. 30). Еще до того, как было продемонстрировано практически использование оптических фильтров в восстановлении изображений, Д. Габор (1949, 1951) математически описал другой способ получения изображения с помощью фотографии. Габор стремился увеличить разрешающую способность электронной микрофотографии. Он предположил возможность интерференции когерентной фоновой волны с волнами, которые преломляются тканью. (Этой цели может служить также отражение волн от темного объекта.) Возникающий в результате интерференции узор сохранял бы и амплитудную и фазовую (взаимодействие по соседству) информацию, которая затем на втором этапе могла бы быть использована для восстановления при освещении источником когерентного света изображения естественной ткани. Габор назвал свою технику «голографией», а сфотографированную картину- «голограммой», потому что она содержала всю информацию, необходимую для восстановления целостного образа. 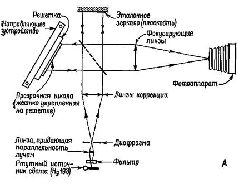 
Голограммы Габора можно построить двумя способами. Волна разделяется с помощью прибора, расщепляющего лучи (то есть полупрозрачного посеребренного зеркала) таким образом, что-одна часть служит в качестве опорной волпы, а другая отражается от объекта, который должен быть сфотографирован (рис. VIII-6 и VIII-7). Затем можно использовать один опорный луч для восстановления изображения. Либо каждая часть расщепленного-луча может быть отражена от различных объектов. Когда при восстановлении изображения один из объектов используется в качестве опорного, другой появляется как образ его «призрака» (фантомное изображение). В этом случае голограмму можно использовать как механизм ассоциативного хранения информации. Постепенно становится очевидным формальное сходство междуголограммами преломления и отражения Габора и различными типами оптических фильтров. Их сходство состоит в том, что-закодированная в каждой из них информация является линейным преобразованием узора световых волн не только по интенсивности света, как в обычной фотографии, но и по их пространственному взаимодействию (пространственной фазе). Наиболее-изученными являются голограммы, у которых эти фазовые отношения могут быть математически выражены посредством преобразований Фурье. Эти голограммы представляют собой особую форму интеграла свертки, который обладает тем свойством, что одно и то же уравнение осуществляет прямую и обратную операции свертки. Таким образом, процесс, выражаемый пространственным преобразованием Фурье, может осуществлять кодирование и последующее декодирование просто в результате возвращения к первоначальному состоянию на некой второй стадии. Все голограммы в целом (рис. VIII-8) обладают рядом интересных свойств, которые делают их потенциально важными для понимания функционирования мозга. Первое – ив данный момент наиболее важное для нас – состоит в том, что информация о каждой точке объекта распределяется по всей голограмме и тем самым делает регистрацию ее устойчивой к разрушениям. Любая малая часть голограммы содержит информацию о всем объекте-оригинале и, следовательно, может восстановить ее. Когда куски голограммы становятся еще меньше, несколько уменьшается ее-разрешающая способность. Когда же для восстановления изображения используются большие части голограммы, уменьшается глубина поля восстановленной картины, то есть сужается зона фокуса. Таким образом, для конкретных целей может быть установлена оптимальная величина голограммы (Leith and Upatniecks, 1965). 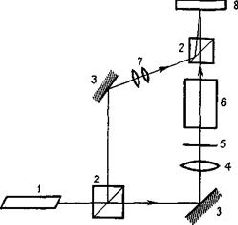
Голограмма обладает фантастической способностью к эффективному (то есть восстановимому) хранению информации. Информация, включенная в соответствующую систему восстановления, может быть непосредственно размещена и точно восстановлена. Плотность хранения информации ограничена только длиной волны когерентного света (чем короче длина волны, тем больше емкость памяти) и зернистостью используемой фотопленки. Более того, одновременно может сохраняться множество различных узоров, особенно когда голограмма наносится на твердый предмет. Каждое изображение хранится по всему твердому телу, хотя каждое из них воспроизводится независимо от других. Как описывают Лейт и Упатниекс (1965),
Обычно в одном кубическом сантиметре голограммы хранится несколько десятков миллиардов бит (единиц измерения) информации. Как заметил П. ван Хирден, если бы мы на протяжении своей жизни каждую секунду запоминали один -бит информации, то для выполнения этой задачи мозг должен был бы каждую секунду совершать около 3 X 10 10 элементарных двоичных операций (нервных импульсов). «Если бы так обстояло дело, то это (прежде всего) было бы невозможно… Однако, столкнувшись с таким парадоксом, постепенно начинаешь понимать… что-оптическое хранение информации и ее обработка могут предоставить в наше распоряжение способ осуществить эту «невозможную» операцию… » (1968, р. 28-29). 
В заключение следует сказать, что оптические системы неединственные системы, которые могут быть объектом топографического процесса. В настоящее время математически описаны и составлены программы для компьютера, «моделирующие» процесс хранения оптической информации. В одной такой программе-интенсивность входного воздействия кодируется размером диска; пространственные фазовые отношения представлены угловой регулировкой щели внутри этого диска (рис. VIII-9). 
Таким образом, голограммы не зависят от физического присутствия «волн», хотя они наиболее полно описываются уравнениями волновой механики. Эта независимость голографии от-наличия физической волны является важным соображением при подходе к проблеме нейронного голографического процесса. Существует достаточно серьезное сомнение в том, что «волны мозга» в том виде, в каком они в настоящее время регистрируются, могут представлять собой субстрат какого-либо узора интерференции, адекватного для переработки информации, хотя они могут указывать на то, что такой процесс имеет место. Конечно, длина регистрируемых волн значительно больше, чем длина волн света, и, следовательно, они могут быть носителями малого количества информации – даже в форме пространственно интерферирующих голографических узоров. Таким образом, предлагаемая далее гипотеза является развитием идеи, высказанной в предыдущих главах о той роли, которую играют в функционировании мозга микроструктуры, образованные медленными потенциалами соединений. Эти микроструктуры могут быть описаны либо в статистических понятиях квантовой теории, либо на языке интегралов свертки и преобразований Фурье, применяемых для описания механической волны. Микроструктуры не меняют своих существенных характеристик из-за того, что мы выбираем тот или другой способ описания. Каждый язык, каждая форма описания имеет свои преимущества. Для объяснения проблем восприятия, особенно проблем формирования образа и фантастической способности узнавания, голографическое описание не имеет себе равных. Поэтому почему же не попытаться применить голографическое описание к процессам мозга? НЕЙРОННЫЙ ГОЛОГРАФИЧЕСКИЙ ПРОЦЕСС Сущность голографической концепции состоит в том, что образы восстанавливаются, когда их представительства в виде систем с распределенной информацией соответствующим образом приво-. дятся в активное состояние. Эти представительства действуют как фильтры или экраны. Фактически, как мы уже отметили, представление о голографической процессе возникает еще при рассмотрении оптических фильтров. В этой связи голография понимается как мгновенная аналоговая кросс-корреляция, осуществляемая в результате сопоставления фильтров. Корреляция в мозгу может иметь место на различных уровнях. На более периферических уровнях возникает корреляция между последовательными конфигурациями, порождаемыми возбуждением рецепторов: остаточные явления, сохранившиеся после адаптации, действующей посредством механизма затухания, создают регистр буферной памяти, которая обновляется текущими входными воздействиями. На более центрально расположенных станциях корреляция влечет за собой более сложное взаимодействие: в любой момент времени входное воздействие коррелирует не только с конфигурацией возбуждения, существующего в любом пункте, но также со структурами возбуждения, прибывающими от других уровней системы. Пример этого вида сложности показан в экспериментах, описанных в гл. VII, где конфигурация изменения потенциалов в зрительной коре определялась не только зрительными стимулами, за которыми наблюдала обезьяна, но также условиями подкрепления и «намерением» осуществить тот или иной тип ответа. Согласно голографической гипотезе, механизм этих корреляций не является следствием ни некоего разобщенного «динамического поля», ни даже изолированных, расщепленных волновых структур. Действительно, рассмотрим еще раз структуру более или менее постоянных организаций кортикальных колонок (или в других участках нервной системы других клеточных ансамблей), возникающих в результате прихода импульсов на нервные синапсы, которые активируют тормозные взаимодействия горизонтальной клетки. При конвергенции таких приходящих динамических структур, исходящих по крайней мере от двух источников, появляются интерференционные картины. Предположим, что оти интерферирующие динамические структуры, образованные классическими постсинаптическими потенциалами, имеют отношение к сознанию. Предположим также, что анализ, проведенный в начале этой главы, правилен и что эта микроструктура медленных потенциалов точно описывается теми уравнениями, которые описывают и голографический процесс, также строящийся на основе интерференционных узоров. Из этого следует вывод, что информация о входных воздействиях распределяется на всем протяжении нейронной системы точно так же, как она распределяется по всему узору физической голограммы. Однако в настоящее время мы еще плохо понимаем, как происходит такой процесс распределения. Следующий обмен мнениями по этой проблеме, проведенный в журнале «Nature», привлекает внимание к некоторым логическим аргументам. 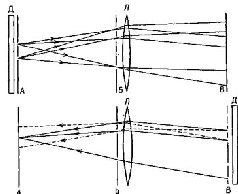
Существование нейронного голографического или сходного с ним процесса не означает, конечно, что входная информация волей-неволей распределяется по всей глубине и поверхности мозга. Информация распределяется только в тех ограниченных областях, где входные воздействия действительно вызывают устойчивые узоры синаптических микроструктур. Более того, для объяснения любого эффекта, развивающегося вслед за специфическим входным воздействием, следует привлечь более локализованные механизмы памяти, описанные в первой части. Однако информация иногда может быть введена в участки, которые распределены по нейронному пространству, и тогда она становится рассеянной. Восстановление того, что более длительное время хранится в памяти, зависит главным образом от повторения данной структуры, которая первоначально вызвала этот процесс сохранения, или ее существенных частей. Эта способность «адресоваться» прямо к содержанию информации безотносительно к ее локализации, которая столь легко достигается в голографическом процессе, устраняет необходимость иметь в мозгу специальные пути или пункты для хранения информации. Какими возможностями располагает нервная система для формирования устойчивых микроструктур соединений? Объяснить хранение информации в течение разного времени можно лишь с помощью какого-то более постоянного свойства протоплазмы. Сильное взаимодействие во времени возникает между входными сигналами, разделенными интервалом в несколько часов (как в случае эффекта Мак-Каллока, когда экспозиция набора цветовых полос оказывает влияние на последующее восприятие цвета) или у некоторых индивидов в несколько дней (как у редко встречающихся людей с настоящей эйдетической способностью). И конечно, для объяснения узнавания и воспроизведения следует принять во внимание более продолжительные процессы взаимодействия. Структурные изменения в таких макромолекулах, как липиды или протеины, и даже длительное анизотропное упорядочение макромолекулярных структур дают основание для следующих рассуждений. Сходные по конфигурации и последовательно возникающие микроструктуры соединений могут порождать остаточный кумулятивный эффект путем упорядочения вначале случайных цепей макромолекул или волокон либо путем увеличения их возбудимости, вследствие чего эта область с большой легкостью отвечает на повторение того же самого возбуждения. Ранние результаты экспериментов, проведенных на культуре ткани сетчатки, которая была исследована с помощью электронного микроскопа, показывают, что такие изменения в молекулярной структуре могут возникать при возбуждении (Sjostrand, 1969). Сходные, хотя и не подтвержденные еще, предположения были сделаны Л. Уайтом (1954) и У. Халстэдом (Katz and Halstead, 1950). Уайт предполагает, что
Конечно, здесь отмечено не все, что свидетельствует о необходимости регистрации волновых форм. Структурные изменения в макромолекулах обладают способностью к восстановлению. Вероятно, более устойчивая запись требует таких механизмов, как настройка «усредняющих циркуляции» в кортикальных колонках и рост, вызванный изменениями в мембранной проницаемости вследствие и в зависимости от этих макромолекулярных изменений. «Фильтр», или «экран», топографических узоров состоит нетолько из решетки макромолекул мембраны, образующих синаптически-дендритную сеть, но также из облегчения всех тенденций формировать образ и создавать выходные динамические структуры нервных импульсов. ПОДТВЕРЖДЕНИЕ ГИПОТЕЗЫ Одна группа данных в пользу голографической гипотезы идет от исследований развития зрительного восприятия у новорожденных. Вопреки широко распространенным представлениям (например, Хебб) константность величины и формы не возникает исключительно из комбинаций следов прошлых и, по-видимому, более элементарных восприятий (например, восприятие длины линий, углов). Предположение Хебба основывалось на результатах экспериментов и клинических наблюдений, когда испытуемые находились в условиях сенсорной депривации. Он считал, что для соответствующего развития рецепторной функции необходим опыт. Однако электрофизиологические эксперименты (воспроизведенные на рисунке VII-12) показывают, что сетчаточные и нейронные механизмы, необходимые для детекции признаков, уже хороша развиты у млекопитающих (кошек) к моменту рождения. Поэтому эффект, наблюдающийся при ограничении сенсорных воздействий, по-видимому, является следствием либо атрофии, возникающей в результате бездействия синапсов, либо активного подавления сенсорного механизма посредством вызванного ненормального функционирования. Как уже отмечалось, у человека возникает аналогичная ситуация. Бауер (1966) проводил эксперименты с шестинедельными младенцами, и эти эксперименты показали, что у них уже наблюдается константность величины ж в значительной степени константность формы. В этих экспериментах предметы помещались таким образом, что их действительная величина, расстояние до них и сетчаточный образ (который вычисляется исходя из действительной величины рассматриваемой фигуры) могли образовывать пары в различных сочетаниях. Использовалось полное и незаконченное изображение фигур-Плоское двухмерное изображение предметов заменяло трехмерные предметы. При обучении младенца различению экспериментатор в качестве подкрепления использовал игру в прятки. Затем производилась проверка, которая состояла в выявлении у младенца заученного ответа при замене одного стимула пары другим. Результаты экспериментов показали, что константность возникает у младенца только тогда, когда он располагает сведениями о расстоянии или когда один элемент фигуры перемещается,, тогда как остальные остаются на прежнем месте. Таким образом, даже у младенцев зрительная система представляет собой врожденную организацию, регистрирующую «такой вид информации, который порождается двигательным и бинокулярным параллаксом». Какой же тип встроенного механизма мог бы регулировать параллакс лучше, чем голографический процесс, который сам конструируется посредством параллактического процесса (эффекта интерференции)? Прямым доказательством того, что в формировании образов участвует параллактический кортикальный механизм, подобный голограмме Фурье, являются результаты экспериментов, проверенных Кемпбеллом и его сотрудниками (Blakemore and Campbell, 1969; Campbell et al. 1968; Campbell, Cooper and Enroth-Cugell, 1969; Campbell and Robson, 1968). Эти исследования показали, что кора головного мозга может настраиваться на восприятие пространственной информации разного диапазона. Эксперименты -были проведены на кошках и на людях. Предъявлялись решетки с различной контрастностью и изучалось влияние их вращения на зрительные вызванные потенциалы (у человека) и на динамические структуры разрядов, возникающих в нейронах зрительной коры (у кошки). Нейроны отвечали на ограниченную полосу пространственной частоты (в четыре октавы), а длительное рассматривание вызывало подавление контрастной чувствительности за пределами соседних частот (рис. VIII-12). Взаимоотношение между этими экспериментальными результатами и голограммой Фурье лучше всего сформулировано самими авторами. 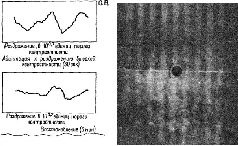 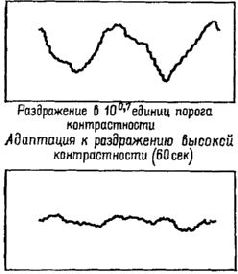
Другая линия доказательств основана на том опыте, который мы все имеем. Можно вспомнить и представить себе множество мельчайших подробностей, когда мы оказываемся в соответствующей обстановке – например, приехав в район, где мы жили много лет назад, мы вспоминаем магазины, расположение дверей, мебель в гостиной, которые еще несколько часов назад казались забытыми навсегда. Какой механизм может действовать лучше, чем ассоциативное припоминание, которое обеспечивается голографическим процессом? Конечно, побудительные импульсы, извлекающие образы из памяти, исходят не от рецепторов. Как я уже отмечал и буду подробно говорить об этом в гл. XVII и XVIII, возбуждение так называемых ассоциативных зон мозга может вызывать во входных каналах динамические структуры возбуждения, аналогичные тем, которые вызываются сенсорными раздражениями. Однако 'Образы, которые предположительно возникают при таком возбуждении, как правило, легко отличить от образов, вызываемых возбуждением рецепторов. Лишь в таких особых условиях, как временная сенсорная депривация, дифференциация внутреннего и" внешнего механизмов формирования образов нарушается, вызывая появление различных иллюзий и галлюцинаций. Однако сам факт, что образы такого типа возникают и формирование их имеет сходство с процессом восприятия, свидетельствует о том,, что восприятие само по себе в значительной степени является реконструктивным. Какой иной процесс, кроме процесса реконструкции образа посредством голографии, может выполнить эту функцию? Главной зоной реконструкции зрительных образов является стриарная область затылочной коры. Известно, что люди, подвергшиеся двустороннему удалению затылочной доли, полностью лишены зрительных образов (Konorski, 1967). Периферическое разрушение не вызывает такого эффекта в слуховой модальности. Прекрасным примером этого может служить Бетховен. Несмотря на периферическое нарушение слуха, он сохранил достаточное воображение, чтобы написать Девятую симфонию и поздние квартеты. Другие данные, согласующиеся с голографической гипотезой,, получены в экспериментах с электрической стимуляцией проекционной области зрительной коры человека (Brindley and Lewin, 1968). Такая стимуляция, если в ней закодирована только интенсивность и не закодированы фазовые отношения, вызывает появление пятен света, выглядящих подобно «звездам в небе» и не похожих на линии или углы. Когда такие пятна возникают в стороне от точки фиксации взора, они становятся слегка удлиненными подобно «зернам риса». Наиболее периферические восприятия выглядят подобно «облаку» и имеют «величину горошин, которые рассматриваются со значительного расстояния». Эти образы появляются на постоянных участках зрительного поля. Однако в результате стимуляции через несколько-отдельных электродов у испытуемого можно вызвать видение-простых зрительных структур. При произвольных движениях глаз воспринимаемые пятна света движутся вместе с глазами; при рефлекторных движениях, вызываемых вестибулярной стимуляцией, воспринимаемые пятна света остаются фиксированными в пространстве. Иногда после очень сильной стимуляции зрительные образы сохраняются в течение одной-двух мипут. Все-эти удивительные наблюдения предполагают существование некоторого механизма, помимо механизма детекции признаков, необходимого для построения весьма сложных зрительных образов,, воспринимаемых нами в повседневной жизни. Еще одна интересная линия доказательств в пользу существования нейронного топографического процесса связана с исследованиями частичных поражений затылочной коры. В соответствии. с моделью простого детектора не следовало бы ожидать каких-либо нарушений вне области скотомы, полученной в результате локальных поражений коры. Но тщательно проведенные исследования Г.-Л. Тейбера и В. С. Баттерсби (Teuber, Battersby and Bender, 1960) показали, что во всей сохранившейся части поля зрения быстрее теряется четкость контуров фигур; снижаются пороги слияния мельканий и нарушается восприятие реального кажущегося движения. Все эти феномены зависят, вероятно, от тормозных взаимодействий, которые представляют собой фильтр, создаваемый эффектами интерференции нейронного голографического процесса. В таком фильтре при большом повреждении следовало бы ожидать скорее изменения качества образов, формируемых оставшейся частью фильтра, чем нарушения самого процесса их формирования. ГРАНИЦЫ ПРИМЕНЕНИЯ ГИПОТЕЗЫ Этот обзор был несколько односторонним. Я не только развивал одну-единственную гипотезу, но и выбрал для изучения одну зрительную модальность и проанализировал обстоятельно только ее пространственные экстенсивные качества. Если же гипотеза лмеет большую сферу применения, она должна с некоторыми изменениями относиться и к другим сенсорным модальностям, к другим сенсорным качествам. С тех пор как исследователи гаптических форм чувствительности (ощупывание) начали серьезно интересоваться проблемой формирования пространственного образа, стало очевидным, что эту гипотезу сравнительно легко, как показал Бекеши, перенести на совместетическую модальность (рис. VIII-3). Модель Бекеши для слухового процесса настолько сходна с нашей, что использование ее нейронного механизма не только возможно, но и дало бы положительные результаты. Это сходство объясняется тем, что высота звука (а следовательно, ж гармония) имеет в нервной системе пространственный код и, таким образом, связана с нейрологическим пространством. Мы слишком мало еще знаем о нейрологических процессах, участвующих во вкусовом и обонятельном восприятиях, чтобы позволить себе высказать нечто большее, чем простое предположение, что дальнейшее исследование не принесет нам данных, которые бы сильно противоречили предложенной здесь модели. Между тем предварительное применение Бекеши (1967) этой гипотезы к вкусовой модальности и Леттвином (Gesteland, Lettvin and Pitts, 1968) к анализу нейронной активности обонятельных луковиц говорит о приемлемости такого подхода. Пока мы не можем сказать ничего больше. Представленная здесь гипотеза иптуитивно отвечает требованиям, которые предъявляет механизм оппонентных процессов в цветовом зрении, как он был предложен Э. Герингом (1920) и экспериментально описан Л. Гурвичем и Д. Джеймсон (I960). Работа Р. де Валуа (De Valois and Jacobs, 1968; рис. VIII-13) с регистрацией в зрительной системе реакции оппонентных нейронов и исследование Г. Светихина (1967) оказались ключевыми для понимания того, каким образом в сетчатке формируется механизм дополнительных цветов. Они подтверждают возможность распространения модели за пределы восприятия структуры раздражителя. Однако да сих пор эти идеи вызывают к себе скептическое отношение, по крайней мере те из них, которые мы рассматриваем здесь и которые еще недавно относились к нейромифологии. 
Более серьезным препятствием для распространения топографической гипотезы является недостаток количественных данных, с помощью которых можно было бы описать способность неврологического процесса обрабатывать информацию. Как мы уже отмечали ранее, имеется серьезное сомнение в том, что «волны»,, которые мы получаем в наших записях, являются субстратом некой организации, создающей узор интерференции, представляющий значение для обработки информации, хотя они могут свидетельствовать о гом, что какой-то сходный процесс действительно имеет место. Длина этих волн такова, что они могут содержать очень малое количество информации – даже принимая форму интер'ферирующих в пространстве голографических: структур. Напротив, развиваемая здесь топографическая гипотеза подчеркивает роль «микроволновых» структур медленных потенциалов, возникающих в синаптических механизмах при функционировании мозга. Однако следует напомнить еще раз,, что эти микроструктуры могут быть описаны либо в статистических квантовых понятиях, либо на языке механической волны в понятиях интегралов свертки и преобразований Фурье. Микроструктуры не меняют своих свойств от выбора того или иного способа описания. Каждый язык, каждая форма описания имеют свои преимущества. Для физической голограммы, получена ли она в результате интерференции световых волн или в результате программирования на компьютере, вычисление количества сохраненной или обработанной информации осуществляется на основе количественного описания пространственных фазовых отношений, определяющих этот процесс. Крайне необходимы эксперименты, которые расшифровывали бы сходные количественные отношения при взаимодействии кортикальных колонок. Основа для таких экспериментов заложена анализом взаимодействия рецеп-торных элементов, выполненным Родиеком и Хартлайном. Несмотря на эти недостатки, многие данные, касающееся функционирования мозга в процессе восприятия и до сих пор казавшиеся парадоксальными, становятся понятными при серьезном анализе голографической аналогии. Это не означает, что все функции мозга сводятся к голографическому процессу или что голографический анализ разрешает все проблемы восприятия. Нейронная голограмма обычно объясняет психологическую функцию формирования образов и механизм распределения памяти в мозгу. Из этого не следует, что память распределена беспорядочно по всему мозгу. Нейронная голограмма объясняет факты, возникающие при разрушении входных систем. Ее распространение посредством экстраполяции на другие системы еще не означает, что системы становятся неотличимыми друг от друга. Даже в процессе формирования образа и, конечно, в узнавании должны играть роль и другие механизмы памяти, помимо тех, которые соответствуют голографической аналогии. Структура этих дополнительных механизмов была детально рассмотрена в гл. VII. Она представляет собой основу нескольких последовательно действующих механизмов. Вместе с тем голографическая аналогия хорошо соответствует системе элементарных логических модулей, набросок которых был дан в первой части книги. Особенно важно, что голографическая гипотеза не опровергает классических нейрофизиологических концепций; она обогащает их тем, что придает особое значение не нервным импульсам аксона, а микроструктуре медленных потенциалов, которая развивается в постсинаптических и дендритных сетях. В то же время голографическая гипотеза обогащает психологию, предоставляя в ее распоряжение правдоподобный механизм для понимания психологических явлений восприятия. Это делает возможным рассмотрение отдельных компонентов психологических функций, которые смешиваются воедино в узких рамках бихевиористского подхода. Распознавание структуры – это сложный процесс, в котором анализ признаков входного сигнала и формирование его центрального представительства являются отдельными этапами. У человека, который наделен нейронной голограммой, эти этапы ведут к формированию образов. Всякая наука ищет объяснительные принципы, и психологическая наука не является исключением в этом отношении. Успех объяснения некоторых загадок восприятия с точки зрения микроструктуры медленных потенциалов соединений и успех самой концепции восприятия как инструмента анализа, расчленяющего поведенческое действие на его функциональные компоненты, заслуживают серьезного внимания. Поэтому в последующих главах мы продолжим рассмотрение этих руководящих принципов применительно к ряду других психологических функций, мотивации и .эмоциям с точки зрения их выражения в субъективных переживаниях. РЕЗЮМЕ Механизм детекции и анализа признаков, осуществляемый нейронами, выполняющими определенные логические операции, недостаточен для объяснения всех феноменов восприятия. Дополнительный механизм содержится в динамических структурах нейронной активности соединений. Происходит суперпозиция, то есть пространственное взаимодействие фазовых отношений между динамическими структурами соседних нервных образований, и такое взаимодействие может лежать в основе способности формирования образов, сходной со свойствами систем, обрабатывающих оптическую информацию, – свойствами голограмм. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||