|
||||
|
|
Глава XIII ДЕЙСТВИЯ МОТОРНАЯ КОРА Четкие, избирательные движения, лежащие в основе поведения, осуществляются моторной корой. Эта кора, расположенная в прецентральной извилине полушарий (рис. XIII-1), связана с базальными ганглиями и мозжечком. Каковы же функции этой моторной коры? 
Уже на протяжении целого столетия идет горячий спор об особенностях организации центральной моторной системы. Некоторые авторы (Woolsey, Chang, 1948) придерживаются точки зрения, согласно которой в моторной зоне существует анатомическое «точка в точку» представительство мышц и даже волокон и моторная кора играет роль клавиатуры для всякой другой церебральной активности и прежде всего для всех произвольных действий. Напротив, другие авторы (например, Phillips, 1965) указывают, что рецептивные поля соседних корковых нейронов охватывают широкий набор мышц, хотя большинство активных мышц, зарегистрированных при раздражении какого-либо одного участка коры, связано лишь с определенным суставом. С таким представлением согласуется тот факт, что электрическое раздражение моторной коры вызывает движения, интегрированные в определенную последовательность мышечных сокращений, и что движения, вызываемые отдельным возбуждением, частично зависят от состояния мозга и положения конечностей, что и отражается на их реакциях при стимуляции. Эти нейрофизиологические данные всегда интерпретировались таким образом, что в моторной зоне коры представлены движения, а не мышцы. Несколько лет назад я еще раз вернулся к этому спору, повторил многие критические эксперименты и накопил новые наблюдения с помощью некоторых дополнительных технических устройств (Pribram, et al., 1955-1956). Результаты этих экспериментов и наблюдений показывают, что основная роль моторных зон коры заключается не в контроле за индивидуальными мышцами или специфическими движениями. Моторная кора, скорее, играет более важную роль в организации действия – действия, определяемого не с точки зрения участвующих в нем мышц, а с точки зрения обеспечения внешнего выражения психологической установки или плана. Это навело меня на мысль, что центральный моторный механизм является в некотором смысле подобием сенсорной системы и что поражение моторной коры вызывает «скотомы в действиях» по аналогии со скотомами в зрительном поле, возникающими при поражении зрительной коры. Появляется все больше данных в пользу этой точки зрения. Вначале я столкнулся со следующим анатомическим парадоксом. Клетка моторной коры получает импульсы через дорсальный та-ламус – структуру ствола мозга, которая расположена дорсально от коры (о чем говорит и само ее название). Следовательно, эта структура аналогична дорсальной части спинного мозга, которая, как показали Белл (1811) и Мажанди (1822), является «сенсорной» частью периферических нервов (рис. XIII-2). Почему же моторная кора должна быть так тесно связана с сенсорными структурами противоположной стороны тела? 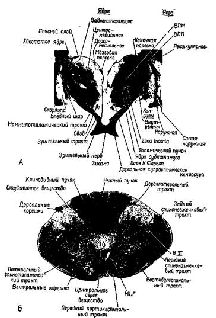
Этот анатомический парадокс особенно ярко проявился в ходе экспериментов, в которых изменения электрических потенциалов вызывались в коре посредством электрической стимуляции периферических нервов. Однажды совершенно случайно было замечено, что такого рода изменения потенциалов возникают в моторной коре. Эти явления слишком противоречили известным фактам, чтобы сразу поверить им. Однако в действительности оказалось, что они были получены и другими авторами, хотя ни один из них не решился их опубликовать и имелось лишь одно упоминание об этом как о возможном артефакте, да и то – в сноске. Между тем наши эксперименты (Malis, Pribram and Kru-ger, 1953, рис. XIII-3) и работы других авторов (Albe-Fessard, 1957; Penfield, Boldrey, 1937) показали, что входные импульсы от периферии действительно достигают моторной коры, что они проходят через дорсальный таламус, а не через уже давно изученные сенсорные зоны коры, такие, как соматосенсорная кора, примыкающая к моторной, и что эти импульсы не зависят от мозжечка. Кроме того, было показано, что входные импульсы возникают не только в тех нервных волокнах, которые иннерви-руют мышцы, но также и в тех, которые связаны исключительно с кожей. 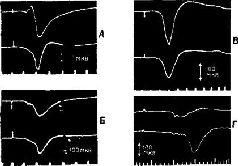 Рис. XIII-З. А - ответы в коре больших полушарий, вызванные стимуляцией седалищного нерва, перед удалением постцентральной коры и мозжечка. Верхняя кривая – ответь в постцентральной коре; нижняя – ответы в прецентральной коре. Время – 10 мсек. Б - ответы сразу после удаления обоих полушарий мозжечка; В - ответы после дополнительного удаления передней доли мозжечка; Г - ответы после дополнительного удаления обеих постцентральных извилин. Заметьте, что в постцентральной области теперь регистрируются только ответы белого вещества мозга (Pribram, Kruger, 1953). Факты, полученные в других наблюдениях и экспериментах, также подтверждают сенсорную природу моторной коры. Обезьяны и человек, у которых была удалена моторная кора, могли выполнять всякого рода движения при соответствующей организации опыта. Инвалиды войны, у которых уже много лет были парализованы руки вследствие поражения моторной коры, находясь в запертой комнате, при пожаре поворачивают дверную ручку парализованной рукой. Когда обезьянам после удаления моторной коры показывали кинофильмы в замедленном темпе, на которых демонстрировались другие обезьяны, неуклюже выполнявшие заученный ряд движений, чтобы открыть щеколду на ящике с орехами, то они без затруднений делали точно такие же движения рукой и пальцами, которые возникают у них при лазании по решетке клетки или при уходе за собой. Затруднены были только некоторые акты, некоторые движения, но эти затруднения не были связаны со специфическим набором необходимых в данном случае движений (Pribram et al., 1955-1956). Каким же образом моторная кора участвует в превращении движения в действие? ОБРАЗ РЕЗУЛЬТАТА Чтобы ответить на вопрос о том, как движение превращается в действие, надо понять, какого рода процессы, связанные с формированием образов, должны происходить в моторной коре и как образ становится мгновенным Образом ожидаемого результата, который содержит в себе всю входящую и выходящую информацию, необходимую для того, чтобы осуществилось следующее звено поведения. Чтобы проверить эту гипотезу, необходимо показать, что, подобно тому как в зрительной коре кодируется незрительная информация (например, образ правой или левой стороны панели, успеха и ошибки), так и моторная кора должна кодировать иную информацию, чем движения. Вероятность того, что такое кодирование должно происходить, велика, так как значительная часть информации, поступающей в моторную кору, исходит из коры мозжечка – структуры, имеющей обширные связи со всеми сенсорными рецепторами. Но каким образом кора мозга становится местом моментального возникновения Образа ожидаемого результата? Каким образом входная информация, поступающая от периферических структур, участвующих в выполнении движения, организуется так, чтобы предвосхитить результат движения, а не просто служить регистратором компонентов движения? Классические эксперименты, выполненные в 30-х годах Н. А. Бернштейном (1966) в Советском Союзе, помогают ответить на этот вопрос. Наряду с последними сведениями о значении гамма-эфферентного контроля над движением теперь начинает проясняться и этот важный и весьма интригующий аспект организации действия. Итак, каким образом организм создает предваряющий Образ, предвосхищение последствий своего поведения? Н. А. Бернштейн проанализировал выполнение таких действий, как ходьба, бег, работа молотком, напильником или печатание на машинке, с помощью кинематографической записи. Испытуемые были одеты в черные костюмы с белыми полосами, чтобы выделить суставы конечностей (рис. XIII-4). В результате снятый фильм представлял собой последовательно развертывающийся рисунок движений, состоящий из отдельных колебаний, которые могли быть математически проанализированы. Бернштейн обнаружил, в частности, что любое ритмическое движение может быть представлено посредством тригонометрической функции и что каждое следующее звено подобного движения может быть предсказано с точностью до нескольких миллиметров по формуле суммы трех или четырех гармонических колебаний – по так называемой тригонометрической сумме Фурье (рис. XIII-5). Из того факта, что эти компоненты движения могут быть описаны в математических терминах и охарактеризованы как «временная голограмма», следует, что именно такого рода Образы, а не предполагаемый клавиатурный механизм, организуют, вероятно, поведение. Следовательно, совокупность явлений, происходящих в моторной коре, можно рассматривать как отражение своего рода голографической трансформации или проекции предшествующих внешних сил, определяющих действие. 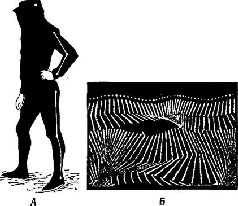
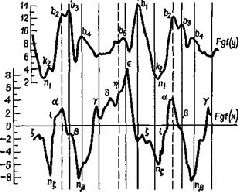
Бернштейн резюмирует эту точку зрения следующим образом:
Как мы вскоре увидим, предположение Бернштейна о том, что в моторной коре представлена топологическая проекция пространства, оказалось ошибочным. На самом деле в моторной коре предстазлены силы, возбуждающие мышечные рецепторы. Однако это не умаляет значения идеи Бернштейна о том, чта в коре больших полушарий кодируются свойства внешней средыа не расположение мышц и суставов. МОТОРНЫЙ МЕХАНИЗМ Как мы уже говорили, наблюдения Бернштейна показывают, что единство действия может быть достигнуто независимо от частных видов движения или количества сокращений каких-либо частных мышц или групп мышц. Действие может осуществляться лишь потому, что движения согласуются с «полем внешних сил», вовлеченных в действие. Как мы уже отмечали при рассмотрении гамма-эфферентной сервосистемы, существенные изменения в этой системе компенсируются указанным механизмом, так что постоянно сохраняется состояние готовности к движению. Таким образом, стабильность состояния достигается за счет специальных приспособлений и компенсаций внешних сил, а представительство в коре больших полушарий этого механизма включает эти параметры. Фактически центральный нервный механизм и состоит в основном, из этих приспособлений и компенсаций; следовательно, центральное представительство должно быть организовано таким образом, чтобы быть «зеркальным отражением» поля внешних сил (аналогичный вывод был сделан и в гл. XII). Явное доказательство того, что нейроны моторной коры – особенно те, от которых идут пути на периферию, – действительно чувствительны к воздействиям, вызывающим мышечные сокращения, было получено в серии других остроумных опытов. В этих экспериментах Э. Эвартс (1967, рис. XIII-6) обучал обезьян работать рычагами, к которым были подвешены различные грузы, чтобы затруднить движения. Когда обезьяны научались двигать рычаг, в момент выполнения задания регистрировалась активность нейронов, расположенных в моторной коре. Опыты показали, что эти нейроны активируются до внешних признаков начала движения (по миографической записи). Тщательный анализ установил, что эта электрическая активность зависит от величины необходимых усилий, а не от величины смещения рычага. Более того, нейроны реагировали главным образом на изменение требуемой силы, хотя не исключено, что некоторая активность связана и с величиной необходимого усилия (рис. XIII-7). 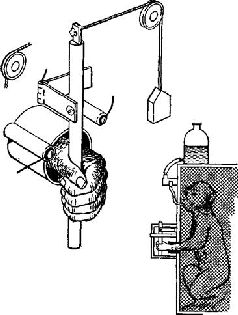
Тот факт, что моторная кора предвосхищает параметры усилия, очень важен. Поскольку рефлексы организуются сервомеханизмами, центральное представительство этих рефлексов составляет не запись длины или напряжения мышц, а запись параметров приспособлений и компенсаций, соответствующих тем изменениям внешних сил, которые вовлечены в движение. Вследствие конвергенции этих процессов в центральном представительстве движений формируется образ не только прежних и текущих изменений внешних сил, воздействующих на систему, но – благодаря быстродействующим расчетам движений, совершающимся в мозжечке, – и тех изменений, которые должны произойти при продолжении двигательной активности. 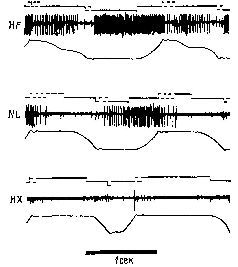
Таким образом, моторную кору можно рассматривать как сенсорную по отношению к действию. Она участвует в пространственной модуляции состояний готовности благодаря своим связям с базальными ганглиями и с механизмом быстродействующего-расчета параметров конечного эффекта действия, осуществляющегося при участии замкнутой мозжечковой системы. Формирование Образа результата зависит – как и всюду в коре – от того, что мы называем «нейронным топографическим представительством», то есть «от в высшей степени избирательного протекания активности в горизонтально расположенных корковых сетях, которые возбуждают и тормозят кортикофугальные нейроны» (Phillips, 1965). То, что это моторное представительство действительно во многих отношениях сходно с тем, которое формируется в сенсорных зонах мозговой коры, может быть установлено при исследовании рецептивных полей нейронов. Результаты этих экспериментов (Welt et al., 1967, табл. XIII-1) показали, что «сенсорная конвергенция в моторной (или сенсорной) коре накладывается на топографически сходные выходные структуры в виде радиальных рядов, диаметр которых, как оказалось, равен 0,1-0,4 мм. Таким образом, нейроны с фиксированными локальными рецептивными полями образуют радиально ориентированную структуру (референтную систему) по отношению к одним и тем же периферическим выходам… Вперемежку с этими клетками, имеющими локальные поля и составляющими 3/4 общей массы клеток, расположены нейроны с широкими, вертикально ориентированными или лабильными полями, которые перекрывают локальные поля» (стр. 285). Здесь, в моторной коре обнаружены явления, которые еще надо найти в зрительной коре (Теперь эти факты уже получены (Spinelli, Pribram and Bridgman, 1971).). Таблица XIII-1 
Остается сделать еще только один шаг в анализе работы моторной коры, а именно указать на то, что уже прекрасно выполнено Родиеком в отношении сетчатки: выяснить, какая именно трансформация точно отражает взаимоотношения между корой и периферией. Итак, контроль над поведением со стороны нервной системы достигается главным образом посредством воздействия на рецеп-торные функции. На рефлекторном уровне чувствительность рецепторов к грузу возбуждает и направляет приспособительный контрпроцесс в сервомеханизме. Сумма таких приспособительных процессов и составляет фоновое тонически устойчивое состояние, на котором возникают новые приспособительные действия. Приспособления широкого диапазона, например изменения позы, контролируются экстрапирамидной системой мозга, в которую входят базальные ганглии и передние структуры мозжечка, тогда как более дискретные движения, такие, как печатание на пишущей машинке, как игра на фортепиано, регулируются быстродействующей неомозжечковой системой осуществляющей экстраполяционные расчеты. Точный механизм этого центрального контроля за движениями пока еще не изучен, однако то, что мы знаем, достаточно, чтобы утверждать, что в этом процессе участвуют явления моделирования периферических сервомеханизмов и что такое моделирование осуществляется посредством механизмов коррекции. Наконец, предлагаемая концепция радикально изменяет наши представления о функциях прецентральной моторной коры. Эта часть мозга, как было показано, является сенсорной по отношению к действиям. Образ конечного результата формируется и непрерывно -мгновенно видоизменяется благодаря нейронным голографическим процессам, так же как и перцептивный Образ. Однако Образ ожидаемого результата создается на основе уже известных из прошлого опыта антиципации тех внешних сил и их изменений, которые требуются для выполнения задания. Эти поля сил,, оказывающие влияние на мышечные рецепторы, становятся параметрами сервомеханизма и непосредственно (через таламус) или опосредствованно (через базальные ганглии и мозжечок) связываются с моторной корой, где они сопоставляются с быстродействующими расчетами мозжечка по предвосхищению результата и последующего звена действия. Когда ход действия становится предсказуем на основании тенденций предшествующих успешных предсказаний, может быть сформирован Образ конечного результата для управления окончательной фазой двигательной активности. Эта модель, которая вытекает из результатов приведенных исследований, существенно отличается от представлений о моторной коре как о клавиатуре, к которой стекается активность остальной части мозга, чтобы разыгрывать мелодию движения. Эта модель, точнее, представление о том, как совершить движение, кодируется в моторной коре и превращается в эффективные действия, акты – внешние выражения мозговых процессов. В следующих главах мы рассмотрим вопрос о том, как эти внешние представительства движений модифицируются в последовательности действий. Последующая часть книги посвящен» исследованию действий, связанных с процессом коммуникации. РЕЗЮМЕ Из ряда мозговых процессов, участвующих в организации поведения, один привлекает особое внимание. Этот процесс включает в себя действие моторного механизма коры больших полушарий и коры мозжечка, формирующего Образ конечного результата с помощью быстродействующих расчетов. Посредством этого-механизма осуществляется экстраполяция последствий непосредственно предшествующих движению изменений во внешнем поле, которые воздействуют на мышечные рецепторы. В этом Образе-конечного результата кодируются, следовательно, внешние явления (например, силы), а не наборы мышечных сокращений. Образ-конечного результата регулирует поведение так же, как термостат регулирует температуру: на диске прибора закодированы пе включение и выключение котла, а лишь пределы допустимо»' температуры. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||