|
||||
|
|
Глава XVIII СИМВОЛЫ ПОБУЖДАЮЩИЕ ЗНАКИ Символы – это побудители к действию. Они обретают смысл на основе их применения в прошлом и состояния организма, использующего их, в настоящем. Этим они отличаются от знаков. Символы как обозначения зависят от контекста, в котором они появляются. Знак  – это роза, и более ничего. Символ – свастика имеет различное значение для еврея и для индуса. Цель данной главы – показать, что символы создаются, когда действия влияют на чувства, на интересы. Фактический материал касается функций лобной коры приматов, той части мозга, которая на протяжении примерно десятилетия, в 40-е и 50-е годы, подвергалась изоляции посредством лоботомии (лейкотомии) в целях излечения больных, страдающих психическими расстройствами. Хотя терапевтическая эффективность этой процедуры остается сомнительной, ее применение вызвало многочисленные нейропсихологические исследования, которые лишь теперь начинают становиться такой отраслью знаний, которая поддается точной интерпретации. ИССЛЕДОВАНИЕ ОТСРОЧЕННЫХ РЕАКЦИЙ Это началось вскоре после первой мировой войны в психологических лабораториях Чикагского университета. Функционализм в области психологии, который был тогда новостью, поставил много методических вопросов. Один из них задал Уолтер Хантер (1913), который был в то время аспирантом: «Возможно ли, что у детей и животных действительно появляются «представления»? И можно ли это доказать?» Он разработал метод, при котором на глазах у ребенка какое-то лакомство, например кусок шоколада, прятали в разных местах при последовательных пробах. При каждой пробе либо ребенка помещали в другое место, либо между ним и местом, где спрятано лакомство, ставили ширму. Несколько минут (или даже часов) спустя ребенка допускали к месту, где было спрятано лакомство, причем это место каждый раз менялось и, таким образом, в противоположность тому, что происходит при различении знаков, теряло всякую определенность. Если ребенок найдет лакомство, значит, у него сохранилось «представление» о спрятанном лакомстве и месте, где оно спрятано, и это представление сохраняется в течение интервала между тем моментом, когда оно было спрятано, и моментом, когда оно было обнаружено. Оказалось, что как дети, так и большинство животных хорошо справлялись с этой задачей, что доказывало их способность к припоминанию. В 30-х годах К. Джекобсен в Йельском университете заинтересовался разработкой методики для определения нарушений, вызванных повреждением лобной коры мозга у приматов. Клинические исследования и лабораторные наблюдения над обезьянами наводили на мысль, что повреждение лобных долей мозга вызывает нарушение каких-то мыслительных процессов. Поэтому Джекобсен счел логичным воспользоваться для изучения последствий повреждения лобной доли вышеупомянутым тестом, который стал для ученых показателем способности к припоминанию. К этому времени было создано несколько вариантов этого теста. В первом варианте приманку на глазах у животного прятали в один из двух одинаковых ящиков, затем закрывали крышку и на несколько секунд (обычно 5-15) ставили ширму между животным и ящиками, после чего убирали ширму и предлагали животному выбрать один из ящиков. Иногда ширма бывала прозрачной, иногда непрозрачной. Только опыты при непрозрачной ширме являлись настоящим тестом на отсроченную реакцию. В одном из вариантов теста применялся «косвенный метод»: какой-то вспомогательный признак, например цветной предмет, на некоторое время показанный над местом, где было спрятано лакомство, обозначал, в каком ящике по прошествии определенного интервала времени можно найти подкрепление. После интервала животному разрешали выбрать один из ящиков, который теперь уже не воспринимался непосредственно. Другой модификацией была методика «отсроченного чередования», в которой сигнал, указывающий место, не применялся вовсе. Подкрепление просто перекладывалось в каждой пробе из одного ящика в другой {рис. XVIII-1). 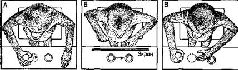
Подход Джекобсена (1928, 1936) к изучению лобной коры оказался удачным: резекция лобного полюса нарушала нормальное выполнение всех вариантов задачи на отсроченную реакцию. Следовательно, от лобных долей каким-то образом зависит способность организма припоминать недавние события. Здесь для нас представляет особый интерес косвенная форма задачи на отсроченную реакцию. Использование этого косвенного метода (выделяющего место) привела некоторых из сотрудников Джекобсена к разработке других задач для исследования способности пользоваться памятными знаками. Наиболее известен прибор под названием «Шимпомат». Автомат выдает фишки, такие, как при игре в покер, и вся «игра» проводится с фишками, которые только позднее и в другом автомате могут быть «обменены» на арахис. Нормальные шимпанзе без затруднений пользуются фишками, этими знаками памяти, но шимпанзе, у которых произведена резекция лобных долей, совершенно не в состоянии оживить следы, ведущие от условного знака к подкреплению (Jacob-sen, Wolfe, Jackson, 1935; рис. XVIII-2). 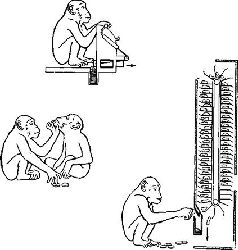
Этот метод обучения получил недавно дальнейшее развитие в другом экспериментальном исследовании, в котором была предпринята попытка научить шимпанзе общению. В гл. XVII описывалась способность обезьяны Уошо пользоваться знаками. Д. Примак (1970) в Калифорнийском университете в Санта Барбара обучал шимпанзе Сару совсем иным методом, чем тот, которым пользовались Гарднеры. Примак применил методы выработки оперантного условного рефлекса для установления того, какой сложности систему условных знаков можно использовать для регуляции поведением Сары. «Шимпомат» уже показал, что шимпанзе будут «работать» за условные знаки. Шимпанзе Примака продемонстрировала не только возможность поведения, зависящего от таких знаков, но также и то, что животное может давать надлежащий ответ и при серийной организации условных знаков (рис. XVIII-3). 
Во всех этих экспериментах суть проблем заключается в том, что условный знак не вызывает единообразного ответа. В зависимости от ситуации (контекста, в котором появляется такой знак) он должен быть понят, перенесен в другое место, опущен в автомат или вручен кому-нибудь, обменен на другой условный знак или на вознаграждение. Либо же, как в первоначальном опыте с отсроченными ответами, знак символизирует вознаграждение, которое должно появиться то в одном, то в другом месте. Для описания этих зависящих от контекста побудительных знаков я буду пользоваться термином «символы». (Это различение терминов согласуется с тем, которое проводит Н. Хомский, 1963, и применяется в данном случае, чтобы указать на то, что истоки правил, определяющих человеческую речь, нужно искать в том, что здесь называется «значимыми» и «символическими» процессами.) Как указывалось в гл. XVII, теперь в нашем распоряжении имеется множество фактов, указывающих на то, что третичные отделы коры, лежащие между классическими сенсорными проекционными зонами в задних отделах мозга, участвуют в различении свободных от контекста знаков. Далее излагаются факты, показывающие, что лобная кора, лежащая кпереди от двигательных зон, участвует в символических процессах, зависящих от контекста. Джекобсен полагал, что процессы, изучавшиеся при помощи этих процедур, должны быть связаны с кратковременной памятью и что эти процессы можно отнести только за счет функции лобных долей мозга. Более поздние исследования показали, что эта гипотеза подтвердилась при двух исключениях. Первое исключение состоит в том, что повреждения хвостатого ядра в области базальных ганглиев (части моторной системы мозга) также-вносят нарушения в выполнение отсроченных реакций. Здесь мы вновь находим доказательство участия двигательных механизмов в осуществлении высших корковых процессов. Второе исключение касается выполнения задач, связанных с отсроченным чередованием; хотя выполнение их и не предполагает использование памятных знаков, повреждение лобных долей нарушает такой вид поведения. Выполнение такой з?дачи страдает также при удалении всех частей лимбической системы (Pribram, Wilson and Connors, 1962). Таким образом, имеются и поведенческие и анатомически» основания для объединения лобных полюсов мозга с лимбиче-скими образованиями (Pribram, 1958, см . рис. XVII-1 и табл-XVII-1). Разрушение этих систем не сказывается на дифферен-цировке знаков, но нарушает выполнение таких задач, как отсроченное чередование (Pribram et al., 1952, 1966; Pribram, Wilson and Concors, 1962), переделка дифференцировки (Pribram, Douglas, Pribram, 1969), поведение приближения или избегания («пассивное» избегание) (McCleary, 1961). При выполнении всех этих задач возникает конфликт в тенденциях ответа, конфликт между установками. Правильный ответ зависит от контекста (то есть от состояния), а контекст изменяется в результате задачи, стоящей перед данным организмом. Таким образом, для получения правильного ответа должна возникнуть внутренняя установка (то есть определенное состояние готовности мозга). Создание такой установки зависит от механизма памяти, включающего систему хранения и воспроизведения установок, возникших в результате периодически повторяющихся функций. (Формальные свойства систем памяти подобного типа были детально описаны Квиллианом, 1967.) Связь лимбических систем по принципу замкнутой петли всегда подчеркивалась анатомами, а это делает эту систему идеальным механизмом, определяющим зависимость действия от контекста (Pribram, 1961; Pribram and Kruger, 1954). Кроме того стоит отметить, что социально обусловленное эмоциональное поведение в очень большой степени зависит от контекста воспринимаемых воздействий. Это заставляет предположить, что значение лимбических формаций для эмоционального поведения обусловлено не только анатомической связью с гипо-таламическими и мезенцефалическими структурами, но также ее собственной организацией по принципу замкнутой петли, поддерживающей раз возникшее возбуждение. Остается доказать (хотя некоторые предварительные данные уже имеются; Fox, et al., 1967; Pribram, 19676), что префронтальная кора оказывает влияние на сигналы лимбической системы примерно так же, как задняя кора участвует в предварительной переработке сенсорных сигналов. Таким образом, связь лобной коры с двигательными механизмами, с одной стороны, и с лимбическими образованиями – с другой, предполагает участие лобной доли в процессе, посредством которого осуществляется связь действия, с одной стороны, с потребностями и эмоциями – с другой. Поэтому способность организма пользоваться побуждающими знаками, или символами, столь зависящая от лобной коры, возникает, по-видимому, в результате этого взаимодействия. ЭМОЦИИ И ПАМЯТЬ Это предположение подтверждается многими фактами. Возможно, наиболее убедительным из них является еще один ряд нейропсихологических парадоксов. Как указывалось в начале этой главы, лоботомия, или лейкотомия, у человека применялась в качестве лечебной процедуры. Применение лоботомии на человеке непосредственно связано с уже описанными экспериментами на обезьянах. Мы видели, что хотя после резекции лобных долей животные оказывались неспособными к выполнению отсроченных реакций, они не проявляли никаких признаков беспокойства в связи с этим. Казалось, что может быть проще, чем ампутировать у больного орган, ответственный за беспокойство? (См. рис. XVIII-4). 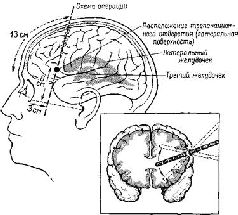
Несмотря на то что такая операция была в известной мере эффективной, исследователи столкнулись с парадоксом, обнаружив, что фактически у больных, перенесших лоботомию, сохранялась способность к решению задач. Пытаясь проанализировать этот парадокс, я провел эксперименты, показавшие, что лобото-мированные обезьяны (а также человек) в действительности проявляли некоторое беспокойство при фрустрации, но это беспокойство было более кратковременным, чем у контрольных испытуемых (Pribram and Fulton, 1954). Этот эксперимент заключается в том, чтобы научить животное правильно дифференцировать сигналы в 100% случаев, после чего время от времени вводились тестовые пробы, не сопровождавшиеся подкреплением. В результате этого возникало беспокойство, отрицательно сказывавшееся на выполнении дальнейших проб, следовавших с равными интервалами. Определялось время, необходимое для достижения установленного критерия (100% правильных ответов). Хотя беспокойство оказывало столь же пагубное влияние после лоботомии, как и до нее, это нарушение продолжалось не так долго. Результаты этого эксперимента привели к предположению, что лобная кора обычно увеличивает устойчивость эффекта неудачи, причем это предположение согласуется с представлением -о важной роли лобной доли приматов в процессах кратковременной памяти. Однако обычные тесты на кратковременную память у людей, перенесших лоботомию, например опыты с припоминанием недавно прочитанных номеров телефонов, не смогли обнаружить никакого отклонения от нормы, хотя болезненные аффективные реакции иа неудачу уменьшились. Но это не исчерпывало наблюдавшихся явлений. В целях излечения некоторых форм эпилепсии нейрохирурги начали вторгаться также и в височную долю человеческого мозга. Казалось-бы, результаты экспериментов с животными свидетельствуют о том, что операции в лимбических областях височной доли приводят к радикальному изменению аффектов. Однако ничего подобного после такой операции у человека не произошло. Вместо этого после резекции можно было наблюдать специфические нарушения памяти. Непосредственная память, как показали тесты с цифрами, оставалась неповрежденной. Так, больной мог припомнить весьма детально свои впечатления перед операцией в сохранял ранее приобретенные перцептивные и двигательные навыки. Однако его способность регистрировать текущие события в долговременной памяти оказалась нарушенной. Стоило прервать серию опытов и возобновить ее через несколько минут или часов, и больной не только не мог припомнить данные им ответы, но даже не имел представления о том, что ему задавали какие-то вопросы или что он вообще принимал участие в эксперименте (Milner, 1958; рис. XVIII-5). 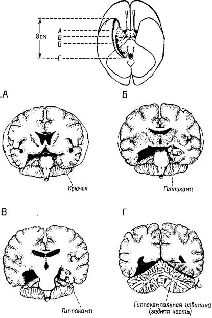
Если бы не этот двойной парадокс, можно было бы легко пренебречь противоречивыми результатами хирургических операций на лобной и височной долях в отношении памяти и эмоций, объясняя их видовыми различиями. Конечно, эти различия представляют собой очевидный фактор, но чем объясняется это удивительное сближение функций памяти и эмоций? Может быть, нарушение функции в основном одинаково у приматов и у человека, но то, что отличает человека, например его владение речью, приводит к существенным различиям между людьми и животными, перенесшими операцию? Возможно, поражение лобной доли сказывается у обезьян во время решения задач, а у человека в тех социальных взаимодействиях, которые протекают без участия речи? Так ли это при лимбических нарушениях? Что касается лобных долей, то на эти вопросы можно дать утвердительный ответ. Сохранная способность больного пользоваться речью позволяет ему решать задачи в опытах с отсроченными реакциями. Однако задачи, слегка усложненные по сравнению с простой отсроченной реакцией, которые, как указывалось, не в состоянии были решить обезьяны, не могли выполнить и люди, перенесшие лоботомию (даже через десять лет после операции), если только эти задачи не сопровождались словесными инструкциями (Poppen, Pribram and Robinson, 1965). Так, больные, которым давалась задача на отсроченную реакцию, говорят,., что они словесно кодируют местоположение спрятанного объекта и таким образом могут припомнить это место после операции, даже если операция происходит после того, как был спрятан объект. Не так легко разрешается парадокс, касающийся лимбическизг повреждений. Прежде всего мы не вполне уверены в том, что при" резекциях височной доли у человека повреждаются только лим-бические структуры. Тяжелая потеря памяти, наблюдаемая у больных, может быть в известной степени воспроизведена у обезьян, когда повреждение захватывает нижнюю латеральную поверхность височной доли, то есть ту часть, которая, как указывалось в предыдущей главе, участвует в опознании и узнавании знаков (Weiskrantz, 1967). Такие обезьяны не в состоянии выработать дифференцировку, возможно, только вследствие разрушения ассоциативной коры височной доли. Когда же резекция ограничивается медиальной частью височной доли, где расположены лимбические структуры (амигдала и гиппокамп), выработка дифференцировки, по крайней мере у обезьян, не нарушается (Pribram, Douglas, and Pribram, 1969). КРАТКОВРЕМЕННАЯ ПАМЯТЬ – КОДИРУЮЩИЙ МЕХАНИЗМ, ОБЕСПЕЧИВАЮЩИЙ СОХРАНЕНИЕ КОНТЕКСТА Предположим, что дело обстоит иначе и рассмотрим гипотезу,, согласно которой фактически существует какая-то единая функция мозга, лежащая в основе как эмоциональных, так и мнести-ческих процессов, причем на эту функцию оказывают влияние лимбические и лобные повреждения. Мы уже видели, что лобные повреждения не препятствуют возникновению эмоционального поведения, лишь сокращая его продолжительность в ситуации фрустрации. Задача, связанная с отсроченной реакцией, также требует длительного сохранения ответа. Может быть, лобная (и лимбическая) резекция просто ускоряет стирание следов памяти? Я подверг эту гипотезу проверке в следующем эксперименте (Pribram, Tubbs, 1967; рис. XVIII-6). Обычно пробы при отсроченном чередовании делаются через равные интервалы (правый ящик – 5 сек, левый ящик – 5 сек; правый ящик – 5 сек, левый ящик – 5 сек и т. д.). Вместо этого я использовал спаренные пробы – правый ящик – 5 сек – левый ящик – 15 сек; правый ящик – 5 сек – левый ящик – 15 сек и т. д. Почти сразу же-обезьяны с лобными повреждениями начали действовать правильно, несмотря на то, что вводились большие (15-секундные) интервалы. Таким образом, гипотеза о более быстром стирании; следов памяти не подтвердилась. 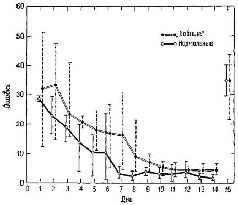
Идея провести этот эксперимент возникла в связи с любимым примером У. Мак-Каллока, который он приводит в качестве доказательства важности кодирования. Когда слова сливаются вместе, как во фразах: угривтине нетихвглине смолавели вдубенет, трудно, а иногда и невозможно найти в них смысл. Но когда вводятся нужные паузы, кодирующие вереницу букв в отдельные куски, сразу же различаются слова: «Угри в тине» и т. д. Задача с чередованием может казаться обезьяне с лобным повреждением слитой в одно целое, ответ интерферирует с предыдущим и последующим. Введение группировки устраняет эту трудность. Организация событий в группы – простой способ обеспечить контекст, необходимый для основных форм кодирования, а также и для более сложного программирования. Возможно, что у человека эта склонность к группировке и организации, к созданию и сохранению контекста, в пределах которого происходят какие-то события, может возрасти до того, что она перейдет границы нормы и явится причиной навязчивости и невроза. Уменьшение этой склонности путем лоботомии на ограниченном участке могло бы в этих случаях привести к успеху! Возможно также, что изменение длительности реакции беспокойства у обезьян, вызванное лоботомией, объясняется тем же самым изменением способности сохранять один и тот же контекст (см. Ornstein, 1969). После операции следы событий сохраняются очень недолго и полностью зависят от контекста воспринимаемых воздействий. Для описания этого аспекта поведения больных, перенесших лоботомию, в клинике употребляется выражение «привязанность к стимулу». Таким образом, эти наблюдения и экспериментальные данные наводят на мысль, что лобная кора участвует в обеспечении и сохранении контекста, временной организации событий. В гл. Хи XV указывалось, что лимбические формации принимают участие в контроле над состоянием мозга. В известном смысле действие лобной коры на лимбический контроль состояний можно сравнить с действием задней «ассоциативной» коры на специфические сенсорные функции. Лобная кора участвует в организации пусковых Образов, создавая зависящие от контекста символы, тогда как задняя кора участвует в организации последовательности перцептивных Образов, обеспечивая сохранение свободных от контекста знаков. В обоих случаях имеет место влияние на Образы. Но это влияние не будет иметь успеха до тех пор, пока не будет предпринято некое действие, некое высшее выражение Образов. Однако нервные структуры, принимающие участие в организации знаков, отличаются от тех, которые участвуют в символических процессах. Перцептивные и двигательные навыки останутся сохранными, несмотря на повреждения лобной и лимбической систем. Напротив, операция кодирования при группировке илвс расчленении событий, необходимая для решения задачи на отсроченное чередование, зависит именно от этих систем. Какие же нервные механизмы принимают в этом участие? САМОРЕГУЛЯЦИЯ ПОВЕДЕНИЯ Пока еще на этот вопрос нельзя дать исчерпывающего ответа. Но некоторые указания относительно такого механизма можно получить, анализируя взаимоотношения между лобной корой и лимбической системой, а также процесс решения задач с отсроченным чередованием и с отсроченной реакцией. Ключ к правильному выполнению теста с чередованием (который сопровождается коррекцией, то есть повторяется до тех пор, пока не будет дан правильный отчет) можно найти в недавно завершенном опыте. Обезьяна должна была составить простой стратегический план, который учитывает контекст, созданный непосредственно предшествовавшим опытом. В этом случае ей очень помогает стратегия «перемещение – ответ». Однако следует отметить, что правильность поведения в любой момент зависит не от событий, происходящих в этот момент в окружающей среде, а от контекста, -созданного памятью (кратковременной). В гл. XV мы выяснили, что амигдала и гиппокамп, входящие в лимбическую систему, регистрируют и оценивают поведение. Поэтому не удивительно, что при повреждении этих структур организм неспособен выполнить задачу, в которой правильность ответа зависит от того, зарегистрирована ли (и оценена ли) предыдущая задача. Чтобы создать стратегический план, связанный с контекстом, контекст должен быть сформирован и сохранен. Однако дефекты выполнения задач на чередование после лобной резекции можно объяснить по-другому. Контекст устанавливается, но организм, очевидно, не знает, как им пользоваться. Мы столкнулись с этой проблемой при объяснении результатов удаления задней «ассоциативной» коры. В этом случае (Bagshaw, Mackworth, Pribram, 1971) запись движений глаз показала, что -обезьяны нормально рассматривали объект и, очевидно, хорошо видели сенсорные сигналы, но не могли действовать на основе того, что они воспринимали. У больных отчетливо проявляется эта неспособность регулировать поведение, когда смысл его зависит от контекста, в котором протекает их деятельность. Клинические наблюдения изобилуют примерами неадекватного поведения. Возможно, наиболее показательным из них является наблюдение, проведенное над одним из больных Г.-Л. Тёйбера (1964). Этот больной регулярно посещал лабораторию по четвергам. Когда один из четвергов совпал с праздником, он, как обычно, отправился в лабораторию, говоря себе на каждом шагу: «Сегодня праздник, мне не нужно идти в -лабораторию, там никого нет». Специальный контекст «праздника», хотя и вербализованный, не оказывал влияния на его поведение. Только когда он дошел до лаборатории и не обнаружил там никого, он вернулся домой, однако так и не сделав вывода о происшедшем. 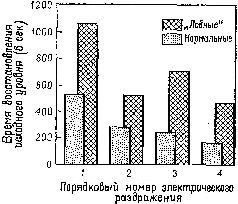
Мы уже много говорили по поводу задачи на чередование. Нарушение, проявляющееся в классической прямой и косвенной формах задач на отсроченную реакцию после резекции лобных долей мозга, объясняет нам еще кое-что относительно взаимоотношений между восприимчивостью к контексту и кратковременной памятью. В этом случае правильность выполнения задания зависит не от контекста, установленного в результате какого-то предшествовавшего действия, а от временного появления сигнала, указывающего на начало пробы. Множество экспериментов показало, что правильная регистрация мозгом этого на мгновение появляющегося знака, способствующая его усвоению путем активной реакции на сигнал, является решающим условием верного выполнения задания (Pribram, 1969b). Что же включает в себя правильная регистрация? В нейрофизиологическом отношении, если нет некоторых изменений в электрической активности мозга, это приведет к ошибке. Именно такие изменения указывают на появление ориентировочной реакции и образуют основу для последующего привыкания. И действительно ориентировочная реакция резко изменяется под влиянием поражений лобных долей как у человека, так и обезьяны (Лурия, Pribram, Хом-ская, 1964). Теперь уже хорошо известно, что имеется ряд индикаторов ориентировочной реакции (Е. Н. Соколов). К ним относится поворот глаз к стимулу, изменение частоты сердцебиений и дыхания, КГР, изменение кровенаполнения -периферических и центральных сосудов (измеряемое при помощи пальцевой и височной плетизмографии) и активация в ЭЭГ. После резекции лобных долей мозга (рис. XVIII-7), так же как после амигдалэктомии, остаются только поведенческие и электроэнцефалографические индикаторы (хотя продолжительность активации в ЭЭГ сокращается). Вместе с тем повторение сигнала в этих случаях уже не вызывает привыкания и поведенческие реакции продолжают сохраняться. Как уже подробно говорилось, это привело к предположению, что ориентировочная реакция состоит по крайней мере из двух отдельных процессов: из компонента настораживания, поиска и отбора стимулов и из их регистрации в- сознании и памяти. Лобная кора и амигдала оказывают влияние на этот процесс регистрации. Когда регистрация нарушается, организм не только утрачивает способность припоминать события, но и становится в гораздо большей степени подверженным ретроактивному и проактивному торможению, обусловленному интерферирующими воздействиями. Люди и обезьяны с лобными повреждениями чувствительны к интерференции, влияющей на процессы кратковременной памяти (Crueninger and Pribram, 1969; рис. XVIII-8). Данные электрофизиологии говорят о том, что лобная кора в норме контролирует интерференцию посредством кортикофугаль-ных влияний на афферентные системы. Электростимуляция лобной коры вызывает результаты, противоположные эффектам стимуляции задней ассоциативной коры (рис. XVIII-9). Раздражение лобных отделов мозга увеличивает избыточность каналов (см. рис. XI-6 и Х1-7; каналы имеют тенденцию в любой момент действовать все вместе. Таким образом, возможность интерференции между следами последовательных сигналов сокращается. Б проводившихся экспериментах длительность вызванного изменения равнялась миллисекундам; ретро- и проактивная интерференция в поведенческих ситуациях занимает значительно большее время. Тем не менее приведенные электрофизиологические данные наводят на мысль, что, по-видимому, в поведенческих ситуациях, в которых наблюдаются эффекты интерференции, лобная кора обеспечивает более длительное сохранение возбуждения, чем нам удавалось достигнуть в эксперименте. 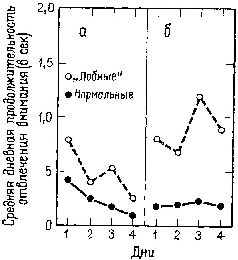

Приведенный анализ опытов с отсроченным чередованием и отсроченными реакциями позволяет предположить, что повреждение лобных долей вызывает нарушение тех процессов мозга, существенным элементом которых является кодирование изменений состояний. Такие процессы свойственны работе кратковременной памяти, предполагающей скорее решения, зависящие от контекста, чем хорошо упроченные действия, не зависящие от контекста, и нарушения их отражаются как на решении задач, так и на эмоциональном поведении. Можно предположить, что подобный дефект сказывается на решении задач потому, что организм не в состоянии регулировать свое поведение на основе событий, вносящих беспокойство и сигнализирующих об изменениях в контексте. Этот дефект проявляется в эмоциональном поведении: организм не справился с контролем, регистрацией и оценкой изменений, непрерывно усложняющих контекст и таким образом увеличивающих беспокойство в настоящем. При повреждении височной доли утрата способности к восприятию контекста проявляется в том, что поток событий не сохраняется в следах и, таким образом, протекает только в настоящем, которое не имеет ни прошлого, ни будущего. Организм становится автоматом, находящимся во власти своих мгновенных состояний, которые он не может регулировать. ПРИПОМИНАНИЕ И СИМВОЛИЧЕСКИЙ ПРОЦЕСС Остается проанализировать связь между кратковременной памятью и символическими процессами. Рассмотрим сначала память. Обычно считают, что процесс сохранения следов происходит примерно так: событие регистрируется в кратковременной памяти, где оно циркулирует в течение некоторого времени, как след в нервных цепях, а затем переводится в более долговременное хранение в молекулярной форме. Этот процесс переноса называется консолидацией следа памяти. Если рассматривать только процессы хранения следов, эту картину можно считать относительно верной. Мы уже отмечали, что привыкание зависит, вероятно, от замкнутой системы с обратной связью, которая уменьшает нервную активность. Ряд таких замкнутых контуров может успешно создать буферный механизм хранения следов в пределах сенсорных систем. Мы также отмечали, что подкрепление может продлить некоторые виды нервной активности, вызывая химические изменения и даже рост глиальной и нервной ткани. Изменения в этих основных механизмах могли бы легко объяснить данные, полученные при экспериментальной проверке гипотезы консолидации. Но анализ процессов памяти только с точки зрения хранения следов не объясняет другие факты, связанные с запоминанием, – узнавание и припоминание. Узнавание предполагает наложение входного сигнала на след, хранящийся в долговременной памяти. Такое наложение во многих случаях может происходить непосредственно, без вмешательства кратковременной памяти, а это значит, что узнавание осуществляется благодаря параллельной обработке информации механизмом памяти, который может восстанавливать опознанный образ даже на основе информации, лишь частично воспроизводящей хранящийся сигнал. Узнавание, однако, не всегда бывает таким непосредственным. В более сложных ситуациях для узнавания может потребоваться некоторое время и оно может зависеть от более медленных процессов выравнивания кортикальных диполей и других процессов. Этот тип узнавания соответствует большинству классических нейрофизиологических точек зрения на процесс памяти. 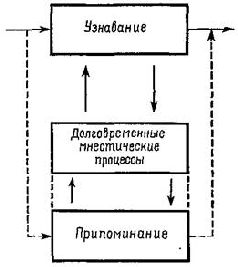
Иной характер имеет процесс припоминания (рис. XVIII-10). Стимулирующее событие запускает механизм кратковременной памяти, который отыскивает в памяти наиболее подходящий материал. В случае узнавания настоящего поиска не происходит – подходящее отыскивается немедленно или очень быстро посредством кросс-корреляции между одновременно возникающими и интерферирующими фронтами волн. Поэтому узнавание «порт-ретно» по отношению к стимулирующему событию и независимо от контекста, в котором оно происходит. Напротив, при припоминании в поиск сходного образца вовлекаются некоторые произвольные «ассоциативные» контекстные связи. Согласно модели, описанной в гл. XVII, лобная стимуляция усиливает торможение побочных связей и, таким образом, повышает связь между модулями памяти. Вовлекаются большие сегменты голографического механизма, фокусное расстояние увеличивается настолько, что внимание сосредоточивается на той или иной детали, то есть концентрируется. Объем и локализация этих усилившихся связей между модулями памяти зависит от предшествующей истории этих модулей, от того, как их следы изменялись непосредственно предшествовавшим опытом. Эта установка выравнивания диполей служит, вероятно, топографическим эталоном, который, будучи активирован неспецифическим входом, запускает посредством опережающего процесса (рис. XVIII-10) реконструкцию «призрака» Образа, ранее ассоциировавшегося с этим эталоном. Возможно, что здесь кроется ключ к пониманию роли лобной коры в процессе припоминания и ее связи с символическими процессами. ЗАИНТЕРЕСОВАННОСТЬ В гл. XVII высказывалось предположение, что регуляция входного сигнала, осуществляемая задней «ассоциативной» корой посредством двигательных структур, служит механизмом придания Образам значимости. В настоящей главе мы привели факты, говорящие о том, что в этом механизме участвуют фронтолимби-ческие отделы мозга, которые часто оказывают противоположное действие по сравнению с задней корой. Таким образом, знаки и символы по-разному участвуют в процессе придания стимулам значения. Знаки извлекают значение посредством избирательного внимания к тем гспектам Образов, которые они обозначают. Символы же извлекают его путем установления контекста, в котором организуются интересы и чувства. Знаки обозначают какую-то часть внешнего мира, какие-то его свойства. Символы выражают то, что зарегистрировано организмом на основе опыта, и его оценку этого опыта – выделение того, что его интересует, к чему он чувствует склонность. Заинтересованность в большой мере заключается в восприимчивости и отзывчивости к изменениям, происходящим в контексте коммуникации. Заинтересованность в ком-то проявляется не столько в каких-то действиях, сколько в том, что это делается в соответствующее время и в соответствующем месте, когда в этом ощущается необходимость. Заинтересованность связана с контекстом и ведет к возникновению адекватных форм поведения. Главным результатом лоботомии является то, что человек становится менее заинтересованным. Мы уже достаточно сказали о различии между знаками и символами; теперь мы обратимся к связи между ними. Эта связь касается человеческого языка, что и является специальной темой следующей главы. РЕЗЮМЕ Символические процессы возникают, по-видимому, благодаря взаимодействию двигательных механизмов с лобной корой и лим-бической системой. Эти части головного мозга характеризуются множеством связей, то есть такой организацией, которая при программировании на ЭВМ ведет к сложным формам коммуникации, связанным с контекстом. Поведение, зависящее от контекста, необходимо для решения тех задач, в котором участвует кратковременная память (припоминание), например, задач на отсроченные реакции и чередование. Оно также необходимо для разнообразных отношений, носящих мотивационно-эмоциональный характер. Участие лобной коры и лимбической системы как в интеллектуальном, так и в эмоциональном поведении зависит, таким образом, от их функции в организации процессов, обеспечивающих связь с контекстом. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Наверх |
||||
|
|
||||